

Carbon and Nitrogen stable isotope changes of mammalian bones from the Niupodong site, Guizhou
Received date: 2024-03-03
Accepted date: 2024-04-09
Online published: 2025-08-07
The recovery of animal prey remains from archaeological sites provides a direct link to human hunting activity and serves as a valuable indicator of paleoenvironmental changes. In this study, we conducted carbon and nitrogen isotope analyses on mammalian bones from the Niupodong site in Guizhou Province to explore the mammalian paleoecological shifts across the first to the fourth cultural phases, with the aim of elucidating the chronological changes in human activities and paleoenvironments spanning from the Upper Paleolithic to Upper Neolithic.
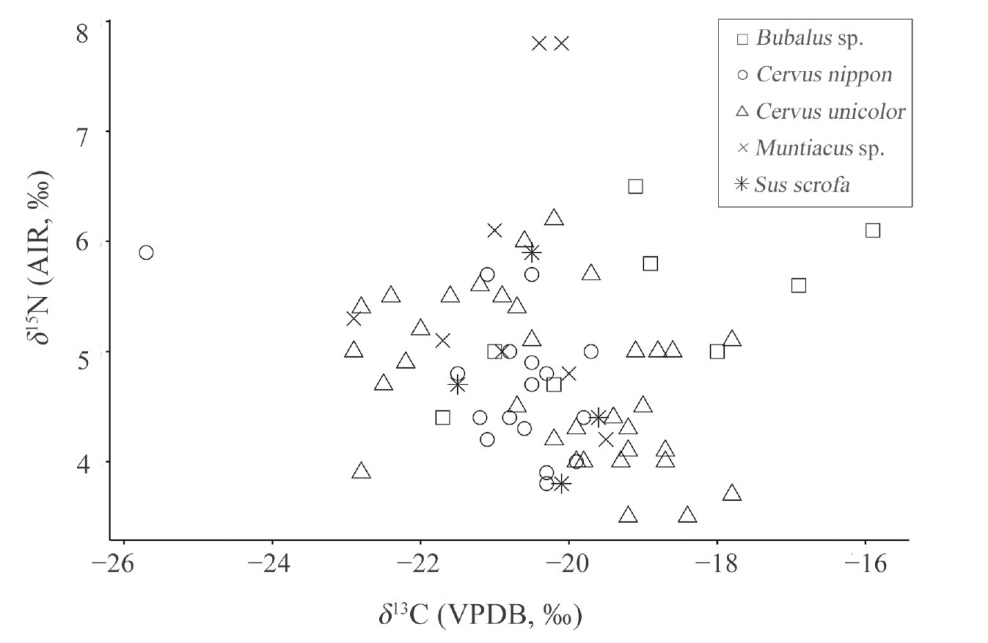
Out of 92 mammalian bone specimens, collagen was successfully extracted from 80, with 72 yielding qualified isotopic data. The mean δ13C and δ15N values of various species including Cervus unicolor (sambar), Cervus nippon (sika deer), Muntiacus sp. (muntjac), Sus scrofa (wild boar), and Bubalus sp. (water buffalo) were −20.3‰±1.5‰ (n=72) and 4.9‰±0.9‰, indicating that these mammals mainly inhabited forest environments dominated by C3 vegetation during the time of ancient human occupation at the Niupodong site. This indicates that these animals were primarily obtained through human hunting, with no apparent signs of domestication.
The range of mammalian δ13C values gradually expanded from Phase 1, potentially indicating an expansion of hunting areas due to increased demand for food resource after Phase 2. Cultural connotation changes were observed during the second and third phases, possibly suggesting advancements in hunting techniques. Coinciding with these cultural shifts, animal bones were highly fragmented at the Niupodong site during the first three phases but became less fragmented during Phase 4. The δ13C range of all the mammals, indicated by the standard deviations of the δ13C values, was narrower during Phase 4 compared to Phase 3, possibly due to the shrinkage of their hunting areas. The standard deviations of δ13C values for sambar specimens were relatively large in the first three periods, peaking in Phase 2 and decreasing to the smallest in Phase 4. This similar trend in data variations among sambar specimens may also reflect changes in hunting intensity.
Based on paleoenvironmental studies, Phase 1 of the Niupodong site, estimated to be approximately 15,000 BP, corresponds to the late glacial period. The second and third phases align roughly with the Holocene climatic optimum, characterized by a strong monsoon signal and a warm, humid climate, while Phase 4 corresponds to a weakening monsoon signal and a trend towards arid, cold conditions. In this study, the δ13C values of water buffalo specimens during the second and third phases were higher than those of Phase 4, exhibiting a C4 signal during Phase 3. Since water buffalo are better indicators of paleoenvironmental conditions compared to cervids, which prefer inhibiting forests, the observed changes are likely associated with large-scale climate variations and shifts in vegetation. Moreover, isotopic data from other animals show minimal temporal changes. This suggests that despite significant paleoenvironmental and paleoclimatic fluctuations across the four cultural phases at the Niupodong site, early humans effectively utilize the animal resources in the surrounding forests, sustaining long-term survival in this region. However, given the gaps between different cultural phases at the Niupodong site, further researches are needed to explore the environmental characteristics of the surrounding areas in the absence of human activity and to determine whether human use of the cave is linked to environmental changes.
MA Jiao , FU Yongxu , CHEN Xianglong , WU Xianzhu , HU Yaowu . Carbon and Nitrogen stable isotope changes of mammalian bones from the Niupodong site, Guizhou[J]. Acta Anthropologica Sinica, 2025 , 44(04) : 688 -700 . DOI: 10.16359/j.1000-3193/AAS.2024.0073
| [1] | 严文明. 中国稻作农业的起源[J]. 农业考古, 1982, 1: 19-31 |
| [2] | He KY, Lu HY, Zhang JP, et al. Prehistoric evolution of the dualistic structure mixed rice and millet farming in China[J]. The Holocene, 2017, 27(12): 1885-1898 |
| [3] | 陈胜前. 中国晚更新世—早全新世过渡期狩猎采集者的适应变迁[J]. 人类学学报, 2006, 25(3): 195-207 |
| [4] | Li HM, Zuo XX, Kang LH, et al. Prehistoric agriculture development in the Yunnan-Guizhou Plateau, southwest China: Archaeobotanical evidence[J]. Science China Earth Sciences, 2016, 59(8): 1562-1573 |
| [5] | Liu RZ, Liu H, Chen SQ. Alternative adaptation strategy during the Paleolithic-Neolithic Transition: potential use of aquatic resources in the Western Middle Yangtze Valley, China[J]. Quaternary, 2020, 3(3): 28 |
| [6] | Roberts P, Perera N, Wedage O, et al. Direct evidence for human reliance on rainforest resources in late Pleistocene Sri Lanka[J]. Science, 2015, 347(6227): 1246-1249 |
| [7] | Gao Y, Dong GH, Yang XY, et al. A review on the spread of prehistoric agriculture from southern China to mainland Southeast Asia[J]. Science China Earth Sciences, 2020, 63(5): 615-625 |
| [8] | Zhang Y, Stiner MC, Dennell R, et al. Zooarchaeological perspectives on the Chinese Early and Late Paleolithic from the Ma’anshan site (Guizhou, South China)[J]. Journal of Archaeological Science, 2010, 37(8): 2066-2077 |
| [9] | Wang XM, Guan Y, Cai HY, et al. Diet breadth and mortality patterns from Laoya Cave: A primary profile of MIS 3/2 hunting strategies in the Yunnan-Guizhou Plateau, southwest China[J]. Science China Earth Sciences, 2016, 59(8): 1642-1651 |
| [10] | 刘晓迪, 王然, 胡耀武. 桂林市甑皮岩与大岩遗址人和动物骨骼的碳氮稳定同位素研究[J]. 考古, 2021, 7: 83-95 |
| [11] | Hartman G, Bar-Yosef O, Brittingham A, et al. Hunted gazelles evidence cooling, but not drying, during the Younger Dryas in the southern Levant[J]. Proceedings of the National Academy of Sciences, 2016, 113(15): 3997-4002 |
| [12] | Naito YI, Hirose M, Belmaker M, et al. Paleoenvironment and human hunting activity during MIS 2 in southern Jordan: Isotope records of prey remains and paleosols[J]. Quaternary Science Reviews, 2022, 282: 107432 |
| [13] | 付永旭, 韦松恒, 文应峰. 贵州平坝县牛坡洞遗址2012-2013年发掘简报[J]. 考古, 2015, (8): 16-36 |
| [14] | 付永旭, 黄超. 贵州牛坡洞遗址新发现的陶器:兼论黔中史前洞穴遗址中的陶器[J]. 南方文物, 2017, (4): 41-50 |
| [15] | 傅宪国, 付永旭, 张兴龙, 等. 贵州贵安新区牛坡洞遗址[J]. 考古, 2017, (7): 3-17 |
| [16] | 王运辅. 贵州贵阳牛坡洞遗址的动物考古学研究[D]. 硕士学位毕业论文, 北京: 中国社会科学院大学研究生院, 2019 |
| [17] | 付永旭. 贵州牛坡洞史前陶器来源初探[J]. 中原文物, 2020, 5: 49-57 |
| [18] | O’Leary MH. Carbon isotope fractionation in plants[J]. Phytochemistry, 1981, 20(4): 553-567 |
| [19] | Farquhar GD, Ehleringer JR, Hubick KT. Carbon isotope discrimination and photosynthesis[J]. Annual review of plant biology, 1989, 40(1): 503-537 |
| [20] | Quade J, Cerling TE, Andrews P, et al. Paleodietary reconstruction of Miocene faunas from Pa?alar, Turkey using stable carbon and oxygen isotopes of fossil tooth enamel[J]. Journal of Human Evolution, 1995, 28(4): 373-384 |
| [21] | Cavagnaro JB. Distribution of C3 and C4 grasses at different altitudes in a temperate arid region of Argentina[J]. Oecologia, 1988, 76(2): 273-277 |
| [22] | Kohn MJ. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate[J]. Proceedings of the National Academy of Sciences, 2010, 107(46): 19691-19695 |
| [23] | Tieszen LL, Boutton TW, Tesdahl KG, et al. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet[J]. Oecologia, 1983, 57(1-2): 32-37 |
| [24] | Cerling TE, Harris JM. Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies[J]. Oecologia, 1999, 120(3): 347-363 |
| [25] | Bocherens H. Preservation of isotopic signals (13C, 15N) in Pleistocene mammals[A]. In: Ambrose SH, Katzenberg MA. Biogeochemical Approaches to Paleodietary Analysis[M]. Boston: Kluwer Academic Publishers, 2002, 65-88 |
| [26] | Sealy JC, Merwe NJVD, Thorp JAL, et al. Nitrogen isotopic ecology in southern Africa: Implications for environmental and dietary tracing[J]. Geochimica et Cosmochimica Acta, 1987, 51(10): 2707-2717 |
| [27] | M?nnel TT, Auerswald K, Schnyder H. Altitudinal gradients of grassland carbon and nitrogen isotope composition are recorded in the hair of grazers[J]. Global Ecology and Biogeography, 2007, 16(5): 583-592 |
| [28] | Bocherens H, Drucker D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems[J]. International Journal of Osteoarchaeology, 2003, 13(1-2): 46-53 |
| [29] | Dalerum F, Bennett NC, Clutton-Brock TH. Longitudinal differences in 15N between mothers and offspring during and after weaning in a small cooperative mammal, the meerkat (Suricata suricatta)[J]. Rapid Communications in Mass Spectrometry, 2007, 21: 1889-1892 |
| [30] | Lohuis TD, Harlow HJ, Beck TDI. Hibernating black bears (Ursus americanus) experience skeletal muscle protein balance during winter anorexia[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2007, 147(1): 20-28 |
| [31] | Richards MP, Hedges REM. Variations in bone collagen δ13C and δ15N values of fauna from Northwest Europe over the last 40 000 years[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2003, 193(2): 261-267 |
| [32] | Stevens RE, Hedges REM. Carbon and nitrogen stable isotope analysis of northwest European horse bone and tooth collagen, 40,000 BP-present: Palaeoclimatic interpretations[J]. Quaternary Science Reviews, 2004, 23(7-8): 977-991 |
| [33] | Fox-Dobbs K, Leonard JA, Koch PL. Pleistocene megafauna from eastern Beringia: Paleoecological and paleoenvironmental interpretations of stable carbon and nitrogen isotope and radiocarbon records[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2008, 261(1-2): 30-46 |
| [34] | Jay M, Richards MP. Diet in the Iron Age cemetery population at Wetwang Slack, East Yorkshire, UK: carbon and nitrogen stable isotope evidence[J]. Journal of Archaeological Science, 2006, 33(5): 653-662 |
| [35] | Price TD, Blitz J, Burton J, et al. Diagenesis in prehistoric bone: problems and solutions[J]. Journal of Archaeological Science, 1992, 19(5): 513-529 |
| [36] | Hedges RE. Bone diagenesis: an overview of processes[J]. Archaeometry, 2002, 44(3): 319-328 |
| [37] | DeNiro MJ. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction[J]. Nature, 1985, 317(6040): 806-809 |
| [38] | Van Klinken GJ. Bone collagen quality indicators for palaeodietary and radiocarbon measurements[J]. Journal of Archaeological Science, 1999, 26(6): 687-695 |
| [39] | MacCullough DR, Takatsuki S, Kaji K. Sika Deer: Biology and Management of Native and Introduced Populations[M]. Tokyo: Springer Tokyo, 2009 |
| [40] | Ahrestani FS, Heitk?nig IMA, Matsubayashi H, et al. Grazing and browsing by large herbivores in South and Southeast Asia[A]. In: Ahrestani FS, Sankaran M. The Ecology of Large Herbivores in South and Southeast Asia[M]. Dordrecht: Springer Netherlands, 2016, 225: 99-120 |
| [41] | McCullough DR, Pei KCJ, Wang Y. Home range, activity patterns, and habitat relations of Reeves’ muntjacs in Taiwan[J]. The Journal of Wildlife Management, 2000, 64(2): 430-441 |
| [42] | 李有恒, 韩德芬. 广西桂林甑皮岩遗址动物群[J]. 古脊椎动物与古人类, 1978, (04): 244-254 |
| [43] | 贵州省文物考古研究所. 贵州开阳打儿窝岩厦遗址试掘简报[J]. 长江文明, 2013, (1): 1-19 |
| [44] | 张弛. 中国新石器化的最初进程[J]. 史前考古, 2024, (1): 1-19 |
| [45] | 陈相龙, 罗运兵, 胡耀武, 等. 青龙泉遗址随葬猪牲的C、N稳定同位素分析[J]. 江汉考古, 2015, (5): 107-115 |
| [46] | 周杉杉. 浙江省余姚田螺山遗址水牛驯化可能性的初步研究:基于C、N稳定同位素食谱分析[D]. 硕士学位毕业论文, 杭州: 浙江大学, 2017 |
| [47] | Leslie DM. Rusa unicolor (Artiodactyla: Cervidae)[J]. Mammalian Species, 2011, 43: 1-30 |
| [48] | 施雅风. 中国全新世大暖期气候与环境[M]. 北京: 海洋出版社, 1992 |
| [49] | Zhou W, Yu X, Jull AJT, et al. High-resolution evidence from southern China of an early Holocene optimum and a mid-Holocene dry event during the Past 18,000 Years[J]. Quaternary Research, 2004, 62(01): 39-48 |
| [50] | Xiao X, Haberle SG, Shen J, et al. Latest Pleistocene and Holocene vegetation and climate history inferred from an alpine lacustrine record, northwestern Yunnan Province, southwestern China[J]. Quaternary Science Reviews, 2014, 86: 35-48 |
| [51] | Dykoski C, Edwards R, Cheng H, et al. A high-resolution, absolute-dated Holocene and deglacial Asian monsoon record from Dongge Cave, China[J]. Earth and Planetary Science Letters, 2005, 233(1-2): 71-86 |
| [52] | 崔琳琳, 胡建芳, 王旭. 末次盛冰期以来中国南方C3/C4植被时空演化及影响机制[J]. 中国科学:地球科学, 2019, 49(8): 1246-1258 |
/
| 〈 |
|
〉 |