

Three-dimensional enamel thickness and bite force of Rudapithecus hungaricus and implications for its dietary adaptation
Received date: 2025-06-20
Revised date: 2025-10-13
Online published: 2026-04-17
Rudapithecus hungaricus, a late Miocene great ape from Rudabánya, Hungary, is a key taxon for understanding the origin, evolution, and dispersal of Homininae. This species has been hypothesized to be capable of processing hard food items. To test this hypothesis, we conducted three-dimensional analyses of enamel thickness (ET) across eight Rudapithecus teeth (including four premolars and four molars) and estimated bite force (BF) of four molars using μCT data. Two enamel thickness indices were measured: Three-dimensional average enamel thickness (3DAET) and the ratio of enamel-thickness to dentine-thickness (3DRED). According to Chai (2018), we measured dentine horn angle (DHA) and cuspal enamel thickness (CET) to estimate BF. Then we compared the results with that of extant great apes (Pan, Gorilla, and Pongo), including 61 ET measurements and 18 BF estimates. Results reveal that the average ET of Rudapithecus is greater in molars than in premolars. While Rudapithecus is broadly akin to Pan with respect to premolar ET, its molar ET shows a pattern more similar to Pongo. Furthermore, the average molar BF of Rudapithecus is 469.66N, which is larger than that of Pan (353.56 N), smaller than that of Gorilla (1023.06 N), and approximately equal to Pongo (499.62 N). Considering a high similarity between Rudapithecus and Pongo regarding molar ET and molar BF, and previous studies have shown that Pongo can process both tough and hard foods, thus our results support the hypothesis that Rudapithecus had the capability of processing hard foods. However, our results do not necessarily suggest that Rudapithecus had an overall tougher and/or harder diet. Which foods a species have actually eaten depends on a variety of factors such as the food processing capabilities, environmental variability, the accessibility of preferred foods, and the competition with other species. Combined with previous dietary researches, it is possible that the functional significance of thick enamel and great molar BF presented in Rudapithecus is to ensure basic energy intake by processing infrequently-exploited exigent mechanically challenging foods (hard and/or tough foods) during periods of fruit scarcity. To advance the understanding of Rudapithecus feeding ecology, future researches may need to incorporate direct dietary evidence derived from various methods such as phytolith, calcium isotope, and dental chipping analyses.
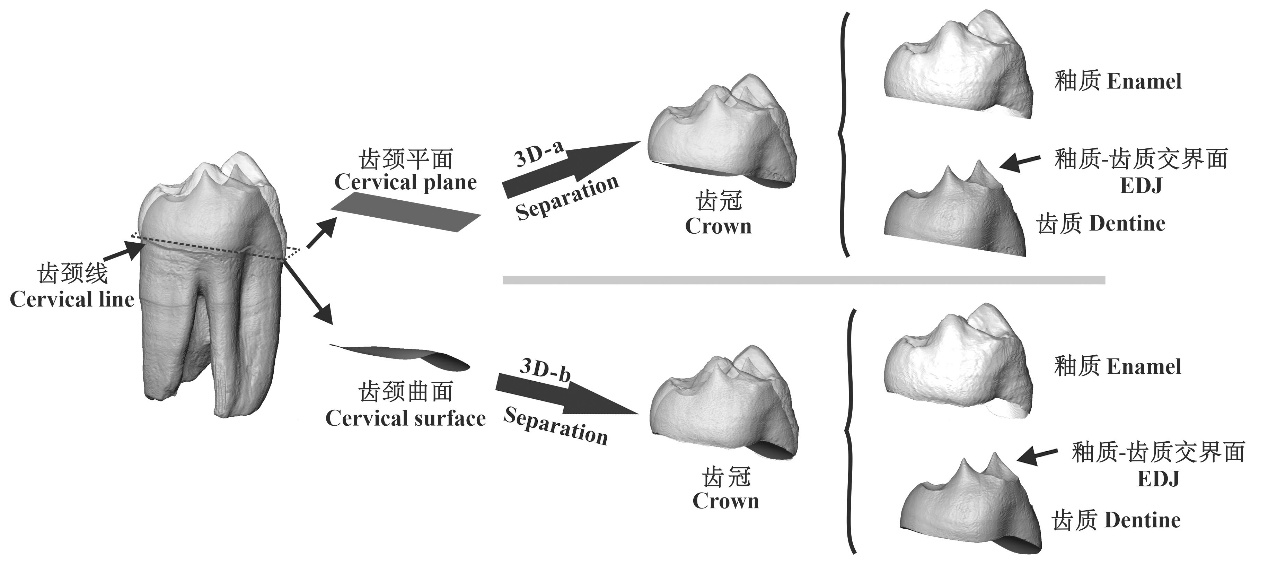
Key words: Rudapithecus hungaricus; 3D enamel thickness; bite force; fallback food
MENG Ziyang , YANG Yuwei , LIAO Wei , WANG Wei , YI Zhixing . Three-dimensional enamel thickness and bite force of Rudapithecus hungaricus and implications for its dietary adaptation[J]. Acta Anthropologica Sinica, 2026 , 45(02) : 258 -267 . DOI: 10.16359/j.1000-3193/AAS.2025.0117
| [1] | Kordos L. The Rudapithecus hungaricus from Rudabánya (Hungary)[J]. Anthropologie, 1991, 95(2-3): 343-362 |
| [2] | Kordos L, Begun DR. A new cranium of Dryopithecus from Rudabánya, Hungary[J]. Journal of Human Evolution, 2001, 41(6): 689-700 |
| [3] | Gunz P, Kozakowski S, Neubauer S, et al. Skull reconstruction of the late Miocene ape Rudapithecus hungaricus from Rudabánya, Hungary[J]. Journal of Human Evolution, 2020, 138: 102687 |
| [4] | Alba DM, Almécija S, DeMiguel D, et al. Miocene small-bodied ape from Eurasia sheds light on hominoid evolution[J]. Science, 2015, 350(6260): aab2625 |
| [5] | Begun DR, Nargolwalla MC, Kordos L. European Miocene hominids and the origin of the African ape and human clade[J]. Evolutionary Anthropology, 2012, 21(1): 10-23 |
| [6] | Kretzoi M. Geschichte der Primaten und der Hominisation[J]. Symposia Biologica Hungarica, 1969, 9: 3-11 |
| [7] | Kordos L, Begun DR. Primates from Rudabánya: Allocation of specimens to individuals, sex and age categories[J]. Journal of Human Evolution, 2001, 40(1): 17-39 |
| [8] | Solà SM, Köhler M. Recent discoveries of Dryopithecus shed new light on evolution of great apes[J]. Nature, 1993, 365(6446): 543-545 |
| [9] | Alba DM, Almécija S, Casanovas-Vilar I, et al. A partial skeleton of the fossil great ape Hispanopithecus laietanus from Can Feu and the mosaic evolution of crown-hominoid positional behaviors[J]. PLOS ONE, 2012, 7(6): e39617 |
| [10] | Kivell TL, Begun DR. late Miocene, New primate carpal bones from Rudabánya (late Miocene, Hungary): Taxonomic and functional implications[J]. Journal of Human Evolution, 2009, 57(6): 697-709 |
| [11] | Ward CV, Hammond AS, Plavcan JM, et al. A late Miocene hominid partial pelvis from Hungary[J]. Journal of Human Evolution, 2019, 136: 102645 |
| [12] | Eastham LC, Feranec RS, Begun DR. Stable isotopes show resource partitioning among the early Late Miocene herbivore community at Rudabánya II: Paleoenvironmental implications for the hominoid, Rudapithecus hungaricus[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 454: 161-174 |
| [13] | Ungar PS. Dental microwear of European Miocene catarrhines: Evidence for diets and tooth use[J]. Journal of Human Evolution, 1996, 31(4): 335-366 |
| [14] | Smith E. A functional analysis of molar morphometrics in living and fossil hominoids using 2-D digitized images[D]. PhD thesis, Toronto: University of Toronto, 1999 |
| [15] | Deane AS, Nargolwalla MC, Kordos L, et al. New evidence for diet and niche partitioning in Rudapithecus and Anapithecus from Rudabánya, Hungary[J]. Journal of Human Evolution, 2013, 65(6): 704-714 |
| [16] | Martin L. Significance of enamel thickness in hominoid evolution[J]. Nature, 1985, 314(6008): 260-263 |
| [17] | Kono RT. Molar enamel thickness and distribution patterns in extant great apes and humans: New insights based on a 3-dimensional whole crown perspective[J]. Anthropological Science, 2004, 112(2): 121-146 |
| [18] | Olejniczak AJ. Micro-computed tomography of primate molars[D]. PhD thesis, New York: Stony Brook University, 2006 |
| [19] | 胡荣, 赵凌霞. 广西化石猩猩牙齿釉质厚度研究[J]. 人类学学报, 2015, 34(3): 404-416 |
| [20] | 张立召, 赵凌霞. 巨猿牙齿釉质厚度及对食性适应与系统演化的意义[J]. 人类学学报, 2013, 32(3): 365-376 |
| [21] | Vogel ER, van Woerden JT, Lucas PW, et al. Functional ecology and evolution of hominoid molar enamel thickness: Pan troglodytes schweinfurthii and Pongo pygmaeus wurmbii[J]. Journal of Human Evolution, 2008, 55(1): 60-74 |
| [22] | McGraw WS, Pampush JD, Daegling DJ. Brief communication: Enamel thickness and durophagy in mangabeys revisited[J]. American Journal of Physical Anthropology, 2012, 147(2): 326-333 |
| [23] | Olejniczak AJ, Smith TM, Skinner MM, et al. Three-dimensional molar enamel distribution and thickness in Australopithecus and Paranthropus[J]. Biology Letters, 2008, 4(4): 406-410 |
| [24] | Smith TM, Tafforeau P, Pouech J, et al. Enamel thickness and dental development in Rudapithecus hungaricus[J]. Journal of Human Evolution, 2019, 136: 102649 |
| [25] | Demes B, Creel N. Bite force, diet, and cranial morphology of fossil hominids[J]. Journal of Human Evolution, 1988, 17(7): 657-670 |
| [26] | Eng CM, Lieberman DE, Zink KD, et al. Bite force and occlusal stress production in hominin evolution[J]. American Journal of Physical Anthropology, 2013, 151(4): 544-557 |
| [27] | Constantino PJ, Lee JJW, Chai H, et al. Tooth chipping can reveal the diet and bite forces of fossil hominins[J]. Biology Letters, 2010, 6(6): 826-829 |
| [28] | Chai H. Dentin horn angle and enamel thickness interactively control tooth resilience and bite force[J]. Acta Biomaterialia, 2018, 75: 279-286 |
| [29] | Chai H. Determining primates bite force from histological tooth sections[J]. American Journal of Physical Anthropology, 2020, 171(4): 683-703 |
| [30] | van Casteren A, Wright E, Kupczik K, et al. Unexpected hard-object feeding in Western lowland gorillas[J]. American Journal of Physical Anthropology, 2019, 170(3): 433-438 |
| [31] | Yi ZX, Liao W, Zanolli C, et al. A robust alternative to assessing three-dimensional relative enamel thickness for the use in taxonomic assessment[J]. American Journal of Physical Anthropology, 2021, 174(3): 555-567 |
| [32] | Yamagiwa J, Basabose AK. Fallback foods and dietary partitioning among Pan and Gorilla[J]. American Journal of Physical Anthropology, 2009, 140(4): 739-750 |
| [33] | Kordos L, Begun DR. Rudabánya: A late Miocene subtropical swamp deposit with evidence of the origin of the African apes and humans[J]. Evolutionary Anthropology, 2002, 11(2): 45-57 |
| [34] | Molnar S. Human tooth wear, tooth function and cultural variability[J]. American Journal of Physical Anthropology, 1971, 34(2): 175-189 |
| [35] | ESRF. Paleontology database[DB]. URL: https://paleo.esrf.eu/.Releasedon:2025-04-17 |
| [36] | Benazzi S, Panetta D, Fornai C, et al. Technical note: Guidelines for the digital computation of 2D and 3D enamel thickness in hominoid teeth[J]. American Journal of Physical Anthropology, 2014, 153(2): 305-313 |
| [37] | Zanolli C, Kullmer O, Kelley J, et al. Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia[J]. Nature Ecology & Evolution, 2019, 3(5): 755-764 |
| [38] | Pan L, Ji XP, Liao W, et al. Premolar enamel thickness and distribution of a Miocene hominid Lufengpithecus hudienensis compared with Pleistocene and extant hominids[J]. Journal of Human Evolution, 2021, 157: 103030 |
| [39] | Nengo I, Tafforeau P, Gilbert CC, et al. New infant cranium from the African Miocene sheds light on ape evolution[J]. Nature, 2017, 548(7666): 169-174 |
| [40] | Olejniczak AJ, Tafforeau P, Feeney RNM, et al. Three-dimensional primate molar enamel thickness[J]. Journal of Human Evolution, 2008, 54(2): 187-195 |
| [41] | Yi ZX, Zanolli C, Liao W, et al. Estimates of absolute crown strength and bite force in the lower postcanine dentition of Gigantopithecus blacki[J]. Journal of Human Evolution, 2023, 175: 103313 |
| [42] | Tafforeau P. Phylogenetic and functional aspects of tooth enamel microstructure and three-dimensional structure of modern and fossil primate molars[D]. PhD thesis, Montpellier: Universite de Montpellier II, 2004 |
| [43] | Smith TM, Kupczik K, Machanda Z, et al. Enamel thickness in Bornean and Sumatran orangutan dentitions[J]. American Journal of Physical Anthropology, 2012, 147(3): 417-426 |
| [44] | Harrison ME, Marshall AJ. Strategies for the use of fallback foods in apes[J]. International Journal of Primatology, 2011, 32(3): 531-565 |
| [45] | Harrison ME. Orang-utan feeding behaviour in Sabangau, Central Kalimantan[D]. PhD thesis, Cambridge: University of Cambridge, 2009 |
| [46] | Wich SA, Utami-Atmoko SS, Setia TM, et al. Dietary and energetic responses of Pongo abelii to fruit availability fluctuations[J]. International Journal of Primatology, 2006, 27(6): 1535-1550 |
| [47] | Rabenold D, Pearson OM. Abrasive, silica phytoliths and the evolution of thick molar enamel in primates, with implications for the diet of Paranthropus boisei[J]. PLOS ONE, 2011, 6(12): e28379 |
| [48] | Tacail T, Le Houedec S, Skulan JL. New frontiers in calcium stable isotope geochemistry: Perspectives in present and past vertebrate biology[J]. Chemical Geology, 2020, 537: 119471 |
| [49] | Towle I, Loch C. Tooth chipping prevalence and patterns in extant primates[J]. American Journal of Physical Anthropology, 2021, 175(1): 292-299 |
/
| 〈 |
|
〉 |