


1 引言
大角鹿(megacerines)是欧亚大陆中纬度地区更新世最常见的哺乳动物之一,在区外仅见于北非地区,该类群在欧亚大陆北部和北非一直延续到全新世[1]。大角鹿类在系统分类上等同于大角鹿族(Megacerini),是鹿亚科(Cervinae)中一支已灭绝的支系[1];古DNA的分析结果显示,爱尔兰大角鹿(Megaloceros giganteus)与黇鹿(Dama dama)的关系最近[2]。大角鹿的一般鉴定特征为角较大,眉枝和主枝或多或少呈掌状,头骨和下颌骨等骨骼发生不同程度的肿厚等[1,3]。欧洲更新世主要是大角鹿属(Megaloceros),我国乃至整个东亚地区则主要是中华大角鹿属(Sinomegaceros),两者的差异主要表现在头骨和角的形态、体型大小以及肢骨特征等方面;中华大角鹿的头骨短宽,下颌骨更肿厚,眉枝更发育,肢骨较短以及体型一般较小[1,4]。我国最为常见的是中更新世的肿骨中华大角鹿(Sinomegaceros pachyosteus, 简称肿骨鹿)和晚更新世的河套中华大角鹿(Sinomegaceros ordosianus, 简称河套大角鹿);前者的化石更为丰富,对其生物学特性也了解较多;而后者的化石材料却很零碎,在已报道过的材料中,发现于陕西榆林雷龙湾(Leilongwan)的一件带角的残破头骨算是最完整的[5]。
2017年,河北师范大学泥河湾考古研究院和历史文化学院考古学系对蔚县北水泉镇一带(属于广义泥河湾盆地范围;图1)开展旧石器时代遗存调查,发现多处石器地点。2018年,对其中文化遗物较丰富的南柏山遗址进行剖面清理和试掘,在中上部地层中发现一件相当完整的大角鹿鹿角化石,除眉枝稍有受压变形以及掌部稍有破损之外,其他部分都保存完好。这件鹿角材料对进一步了解河套大角鹿的生物学特性至关重要,本文将对其形态分类及年代地层等方面的初步研究成果作一报道。
图1
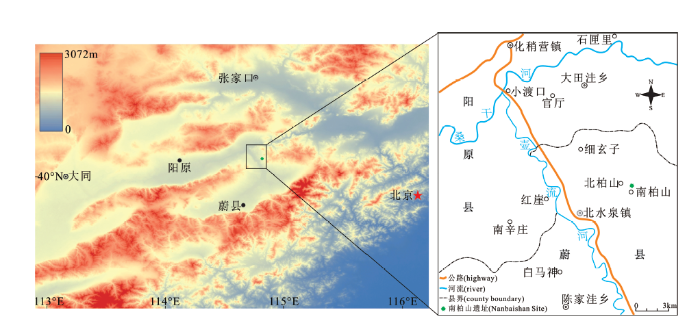
图1
南柏山遗址地理位置图
注. 数字高程模型(DEM)数据来源于我国地理空间数据云平台的GDEMV2 30M分辨率数字高程数据,然后经软件ArcGIS 10.8处理
Fig.1
Location map of Nanbaishan site
2 南柏山遗址地质背景
图2

结合地貌部位、沉积物特征和石制品初步分析判断,南柏山遗址应属旧石器时代中期,地质时代为晚更新世。
3 材料与方法
大角鹿化石出自发掘区东北部的第4a层,是一件几乎完整的自然脱落的右侧鹿角(编号:18NBS(4): 006)。由于地层总体由西北向东南倾斜,鹿角的扁平延展面也向南部倾斜,长轴方向200°,倾向205°,倾角约28°。鹿角风化较重,随即进行了现场加固保护,打石膏包整体取起。
使用Artec Spider 3D扫描仪对化石进行了全方位扫描和三维重建,MeshLab软件进行渲染。
4 系统古生物学
哺乳动物纲Mammalia Linnaeus, 1758
偶蹄目Artiodactyla Owen, 1848
反刍亚目Ruminantia Scopoli, 1777
鹿科Cervidae Gray, 1821
鹿亚科Cervinae Goldfuss, 1820
大角鹿族Megacerini Viret, 1961
中华大角鹿属 SinomegacerosDietrich, 1933
河套中华大角鹿 Sinomegaceros ordosianus(Young, 1932)
图3
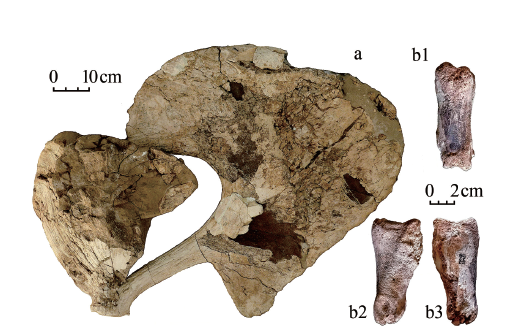
图3
河北蔚县南柏山遗址的河套大角鹿化石
a. 18NBS(4):006,右角right antler (内侧视medial view);b. 17NBS(4):002,第一趾节骨first phalanx (b1.前视anterior view; b2.中侧视interdigital view; b3.外侧视lateral view)
Fig.3
New remains of Sinomegaceros ordosianus from Nanbaishan, Yüxian of Hebei
表1 河套大角鹿鹿角测量及相关对比(mm)
Tab.1
| 属种taxon | 河套大角鹿S. ordosianus | 肿骨鹿S. pachyosteus | 扁角鹿S. flabellatus | 洛川大角鹿S. luochuanensis | ||||
|---|---|---|---|---|---|---|---|---|
| 地点Locality | 南柏山 Nanbai- shan | 丁村 Dingcun[35] | 周家油坊 Zhoujia- youfang[36] | 闫家岗 Yanjia- gang[33] | 灵井Lingjing[37] | 周口店 ZKD 1[11] | 周口店 ZKD 13[5] | 陕西洛川[26] Luochuan |
| 角总长度L(Antler) | 930 | - | - | - | - | 260-576 | 950-1050 | 530 |
| 角环周长C (Burr) | 310 | - | - | - | - | - | - | - |
| 角环前后径d(Antero-posterior of burr) | 104.7 | 78.0-99.8 | 62.0-94.8 | 41.1-91.5 | 103.3 | 74-94 | - | 65 |
| 角环内外径d(Medial-lateral of burr) | 96.4 | 74.0-99.0 | 68.2-95.5 | 49.0-97.1 | 98.2 | - | - | 54 |
| 眉枝最大高Hmax(Brow tine) | 480 | - | - | - | - | - | 350-530 | 174 |
| 眉枝最大宽bmax(Brow tine) | 510 | - | - | - | - | - | 250-300 | 260 |
| 眉枝基部宽b(Base of brow tine) | 114.6 | 74.5-88.0 | 59.5-92.7 | 43.3-97.2 | - | - | - | - |
| 主枝角干周长C (Shaft) | 193 | - | - | - | - | - | - | - |
| 主枝角干前后径d(Antero-posterior of shaft) | 59.8 | 48.0-51.1 | 50.4-87.0 | 32.5-70.5 | 53.7 | 46-94 | - | 56 |
| 主枝角干上下(横)径d(Superoinferior of shaft) | 56.7 | 53.3-69.0 | 51.5-95.0 | 22.5-56.9 | 45.3 | - | - | 42 |
| 主枝角干长L(Shaft) | 270 | - | - | - | - | - | - | 120 |
| 主枝掌部最大长Lmax( Palmation) | 526.8 | - | - | - | - | - | - | - |
| 主枝掌部最大宽bmax(Palmation) | 670 | - | - | - | - | - | 450 | 400 |
产地:河北蔚县南柏山遗址。
层位:晚更新世。
描述:该件鹿角标本最大长近1 m,属于大型鹿角;除角环和掌部稍有破损外,其余部分保存状况完好。
角环:角环(或者角节)是围绕角干基部一周的嵴状隆起,如同一个紧箍环;除过腹侧面发育较弱之外,总体来说角环还是十分发育的,尤其是在眉枝基部处。角环位置略高于角干基底面。从底面观察,角环及其所包围的角封(seal)轮廓近椭圆形, 角封总体圆凸,但中心部位凹陷(图4: d)。角环上未见滋养孔。
图4
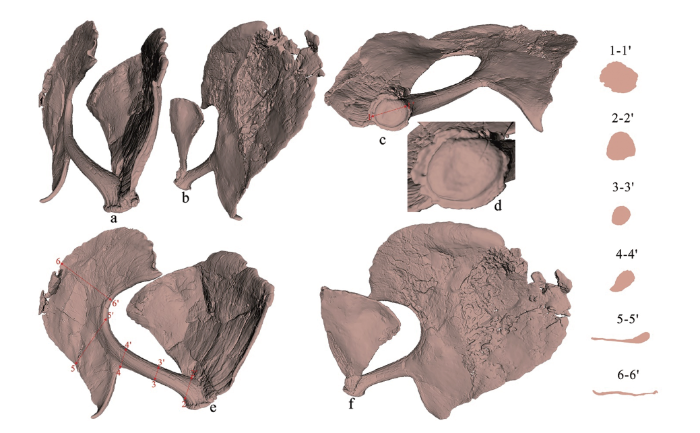
图4
河套大角鹿右角化石的激光扫描三维重建
a.前侧视(anterior view); b.后侧视(posterior view); c-d.底端视(basal view); e.外侧视(lateral view); f.内侧视(medial view)
(鉴于标本的三维虚拟图像是在角度较大的透视视角下输出的,所以不同部位的比例差别很大,故此图没有标注比例尺)
Fig.4
3D-reconstruction of antler fossil of Sinomegaceros ordosianus
眉枝:眉枝基部着生在主枝内侧的角环之上,向上逐渐以放射状变宽,整体呈扇片状;眉枝边缘整齐,无任何指突类结构,但有波浪状突起。背面布满放射状纵纹,腹面的纹饰较弱。眉枝在前内侧与主干在角环处就分开,而在后内侧面与主干的分叉位置较高;该特征与雷龙湾的河套大角鹿标本一致。就目前的保存状况而言,眉枝的延展面与主枝掌状部分的基本一致或以小角度相交;由于眉枝中部的纵向折断,使后半段的走向发生错位,所以难以正确判断整个眉枝的伸展方向。
主枝:分为角干和掌部。角干部分靠基部明显向下折弯,主体呈向前稍凹的弓形,“S”形不甚明显;角干基部最粗壮,中下部横截面近圆形,越向远端(靠近掌状部分)越扁;角干表面布满纵纹,但无棱脊状结构。掌部很大,整体呈银杏叶状,但不完全对称,在接近眉枝部分,即靠前侧,形成一很大的宽缓的切迹,而下缘几乎与角干形成直角;前边缘较盾圆,后边缘较陡直。除过两个近端的尖角和一些波浪状的小突起之外,掌部边缘上几乎无其他指突。掌部基本在同一平面延展,与头骨矢状面斜交;但在两个尖角处稍微向上卷曲。掌部的厚度是从角干向远端逐渐辐射状变薄。
图5

图5
河套大角鹿复原图(许勇绘制)
Fig.5
Reconstruction of Sinomegaceros ordosianus (Drawn by Xu Yong)
关于大角鹿的系统发育尚存不同观点[1,3,4,18⇓-20],其中Vislobokova在2013年详细论述了大角鹿族(Megacerini)、鹿族(Cervini)以及上新鹿族(Pliocervini)三者之间的形态区别,并将大角鹿族进一步划分为10属25种[1]。总的来说,下颌骨肿厚程度[5,21,22]和眉枝形态[3,4]经常被作为大角鹿系统分类的重要特征,不过下颌骨是否可以作为分类的重要依据尚存争议,此外关于大角鹿下颌骨肿厚指数的计算,存在若干不同方法[23]。Azzaroli根据眉枝形态将大角鹿类分成两组:一组以爱尔兰大角鹿为代表,眉枝呈掌状;另一组以直角大角鹿(Megaloceros verticornis)为代表,眉枝横截面为圆形[3]。从掌状眉枝来看,其所在平面与头骨矢状面之间的关系是区分不同属种的一个重要特征,大致可以分为三种类型:垂直于头骨矢状面、与头骨矢状面斜交、平行于头骨矢状面[4]。此外,van der Made和Tong[4]将大角鹿属(Megaloceros)和中华大角鹿属(Sinomegeceros)作为两个独立的属对待,二者之间角的形态区别主要在于3个方面:大角鹿属的眉枝发育相对较弱;大角鹿属存在第二枝甚或第三枝,而中华大角鹿无;大角鹿属的角展开长度一般远大于中华大角鹿。基于此来判断,出土于南柏山遗址的这件鹿角标本毫无疑问属于中华大角鹿。
截至目前,中华大角鹿属在不同地区共报道过以下几种或亚种(图6):中亚地区报道的塔吉克中华大角鹿(Sinomegaceros tadzhikistanis)[1];中国报道的公王岭中华大角鹿(Sinomegaceros konwanlinensis)[24]、扁角中华大角鹿(Sinomegaceros flabellatus)[25]、肿骨鹿[11]、洛川中华大角鹿(Sinomegaceros luochuanensis)[26]、桑干河中华大角鹿[10]、河套大角鹿[5]、河套大角鹿门头沟亚种(Sinomegaceros ordosianus mentougouensis)[27]、包头中华大角鹿(Sinomegaceros baotouensis)[27]和杨氏中华大角鹿(Sinomegaceros youngi)[28];朝鲜半岛报道过祥原中华大角鹿(Sinomegaceros sangwonensis)和扁角中华大角鹿[29];日本报道过矢部中华大角鹿(Sinomegaceros yabei)[30]。其中刘嘉龙和金福全报道的杨氏中华大角鹿[28]与肿骨鹿相比,并没有什么特别之处,属于同物异名[31]。
图6
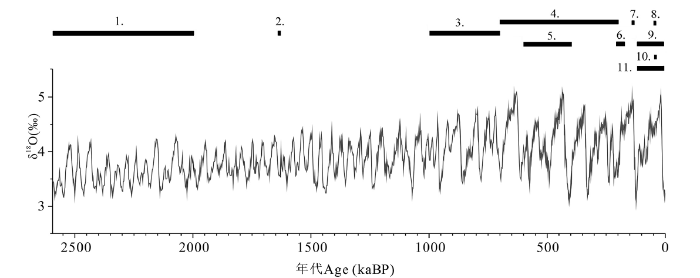
图6
中华大角鹿的地史分布
气候曲线数据源自LR04,参考文献[38]
1.塔吉克中华大角鹿Sinomegaceros tadzhikistanis; 2.公王岭中华大角鹿Sinomegaceros konwanlinensis; 3.扁角中华大角鹿Sinomegaceros flabellatus; 4.肿骨中华大角鹿Sinomegaceros pachyosteus; 5.祥原中华大角鹿Sinomegaceros sangwonensis; 6.洛川中华大角鹿Sinomegaceros luochuanensis; 7.桑干河中华大角鹿Sinomegaceros sangganhoensis; 8.包头中华大角鹿Sinomegaceros baotouensis; 9.河套大角鹿Sinomegaceros ordosianus; 10.河套大角鹿门头沟亚种Sinomegaceros ordosianus mentougouensis; 11.矢部中华大角鹿Sinomegaceros yabei
Fig.6
Geological distribution of the genus Sinomegaceros
The data of the climate curve was adopted from LR04 [38]
与塔吉克中华大角鹿相比,该种的眉枝和角干末端还未发育成典型的掌状形态[1]。与公王岭中华大角鹿相比,该种的眉枝分叉,角干较长,主枝掌部较小且有3-4个指突[24]。与出土于周口店地区的肿骨鹿和扁角鹿相比,这两个种的眉枝所在平面与头骨矢状面近似垂直,且主枝掌部上有多个大小不一的指突、主枝角干短且较扁,下颌骨十分肿厚[5,11]。肿骨鹿的眉枝与主枝掌面近于平行(Young, 1932: Pl. XIII-1和1a)[11],这一特征与蔚县南柏山标本十分一致,只是前者的主枝角干较短而已。其实周口店1地点的大角鹿鹿角化石相当复杂,依据眉枝大小、主枝掌部愈合程度及主枝角干的长短,将其分为3种类型[11],但无一有完整化石记录,因为共生的头骨和下颌骨并未见明显分异,因此推测这3种类型很可能代表大角鹿的不同年龄阶段。扁角鹿的角可分为类型I(主枝角干较长且眉枝面与主枝掌面夹角45°)和类型II(主枝角干较短且眉枝面与主枝掌面近乎平行);扁角鹿的最大特征是眉枝呈三角形,且其延伸面与头骨矢状面垂直,左右眉枝接触或有重叠[5]。值得说明的是,周口店第3地点出土的大角鹿未定种的眉枝与主枝掌部愈合[32]。周口店应当是中华大角鹿化石最丰富和物种多样性最高的地区。尽管周口店地区的大角鹿变异很大[11],但就已报道的大角鹿化石而言(第3地点除外),其共有特征是眉枝着生位置更靠角环前侧位置,而与着生位置更靠角环内侧的河套大角鹿有别。
河套大角鹿广泛分布于我国东北、华北和西北地区的晚更新世地层[33⇓⇓⇓-37],浙江舟山海底出土的标本是该种目前最靠南的记录[8]。河套大角鹿的分布范围非常广,但保存状况好的鹿角标本非常少。出土于陕西榆林雷龙湾的河套大角鹿的鹿角标本[5]保存相对完整,出土于南柏山遗址的鹿角标本的眉枝位置及走向与河套中华大角鹿基本一致,不过两者的主枝延伸方向有较大不同;出土于雷龙湾的河套大角鹿的主枝向两侧延伸,掌状部分所在平面接近水平,而南柏山遗址出土的鹿角主枝则向侧后方延伸,其所在平面与水平面斜交,这可能属于种内变异或鹿角处于不同的发育阶段所致;此外,南柏山遗址的角干“S”形不显著,并且横截面浑圆而无棱角。南柏山遗址鹿角标本的相关测量数据与丁村[35]、周家油坊[36]、闫家岗[33]以及许昌灵井[37]等遗址出土的河套大角鹿近似(表1),但眉枝基部更宽一些。与河套大角鹿门头沟亚种相比,该亚种的眉枝呈薄片状,但外卷成筒形,即眉枝生长是先向上、再向外、后向下直抵主枝角干,这一特征是该亚种独有的,其他属种中缺失;主枝角干细长,上侧面有一明显棱脊,基部横截面呈三角形;主枝掌部特征不明[27]。最近,在萨拉乌苏新发现了河套大角鹿主枝掌部的残片1) (1) 内蒙古博物院李虹通信告知。),其基本特征与本文所记述新材料极为相似。
目前关于大角鹿类的起源主要有两种观点,一种观点认为大角鹿类由早更新世的真枝角鹿(Eucladoceros)演化而来[27],另一种观点认为大角鹿类是一个独立的演化支系,大角鹿族和鹿族共同起源于晚中新世的祖鹿(Cervavitus),随后二者平行演化[1]。而关于中华大角鹿这个支系的起源相对更加明朗一些,目前的化石证据显示中华大角鹿起源于早更新世的中亚地区[1],在中-晚更新世期间,该类群在东北亚地区繁荣发展,目前还未在全新世地层发现过中华大角鹿。根据目前中华大角鹿的地史分布,我们可以初步推断中华大角鹿在中亚起源后,向东扩散到中国西北和华北地区,然后以华北地区为中心,向周边扩散,经由朝鲜半岛扩散到日本,期间演化出具有不同角型的种。
大角鹿是我国北方地区古人类遗址中最常见动物之一,是进行史前遗址间年代对比的重要证据。此外,大角鹿也承载了一定的古人类活动信息,不少旧石器考古学家将鹿角作为工具来打击和加工石器,来推测史前时期古人类使用角器可能进行的生产活动。肿骨鹿和葛氏斑鹿是周口店北京猿人地点最常见的大型哺乳动物,前者的化石材料相当丰富,因此大角鹿与古人类的关系受到学者们的持续关注和研究[39⇓-41]。有学者推测,北京猿人在夏末和初秋更多地狩猎斑鹿,而在冬季则主要狩猎肿骨鹿,北京猿人是猎鹿人[39,42];此外,北京猿人遗址中还发现大量烧烤过的鹿角碎片,这毫无疑问是与古人类活动有关。目前看来,有关北京猿人遗址中动物化石的来源问题尚未取得统一认识。安徽和县人遗址大量的葛氏斑鹿角化石,据推测可能也被古人类所利用[6]。闫家岗遗址的河套大角鹿角上有人工砍痕和非自然的刻划纹[33]。我国发现的唯一旧石器时代鱼叉—辽宁海城小孤山遗址中的鱼叉,其原料也可能是取自鹿角[43]。出土于周口店山顶洞的我国最早的打磨工具和未解刻画图案,也是以鹿角为原料和载体[44]。
鹿类动物化石有很好的时代指示意义。泥河湾盆地的鹿类动物化石较为丰富,而鹿类动物是良好的环境指示分子,常被用来推断古气候环境[49]。泥河湾盆地从中更新世到晚更新世,河套大角鹿替代了桑干河大角鹿,前者掌部明显比后者发育,暗示晚更新世的泥河湾盆地可能比中更新世更加开阔。
5 含河套大角鹿化石的地层时代
发掘同时,在鹿角化石东西两侧各采集一个光释光样品(样品编号分别为18NBS-OSL-01、18NBS-OSL-02)。样品是用直径5 cm、长20 cm的不锈钢管垂直打入地层剖面中采取,随后避光保存。由于地层是倾斜的,所以同一层位中的两个样品埋藏不同,二者埋深相差26 cm,样品18NBS-OSL-02位置略低于化石。样品的等效剂量测定于河北师范大学释光测年实验室完成,在实验室暗室中提取了石英(4-11µm)细颗粒用于等效剂量测量,测量方法为单片再生剂量法。在样品剂量率的测量与计算中,样品中U、Th、K含量的测定采用ICP-MS法于长安大学地球科学与资源学院完成,样品的含水量设定为15%±5%,结合样品的放射性元素含量和含水量,以及埋深、海拔、经纬度等参数,可利用剂量率及年龄计算器DRAC(Dose Rate and Age Calculator)v 1.2计算出样品的剂量率值。最终测量结果列于下表,两个样品的光释光年代分别是109.9±4.2 ka和108.1±4.6 ka,二者的年龄在误差范围内保持一致,说明大角鹿化石所在地层的年代大约为距今11万年前,相当于深海氧同位素5d阶段。
表2 含大角鹿地层中石英样品的光释光测年结果
Tab.2
| 样品编号Sample | 铀含量 U(/106) | 钍含量 Th(/106) | 钾含量 K(%) | 水含量 H2O(%) | 剂量率 Dose rate (Gy/ka) | 测片数Number of aliquots | 等效剂量 Equivalent dose(Gy) | 年龄 Age (kaBP) |
|---|---|---|---|---|---|---|---|---|
| 18NBS-OSL-01 (东侧East) | 1.93±0.04 | 13.07±0.16 | 2.06±0.04 | 15±5 | 3.70±0.13 | 7 | 406.20±6.56 | 109.9±4.2 |
| 18NBS-OSL-02 (西侧West) | 2.06±0.05 | 9.10±0.06 | 2.10±0.07 | 15±5 | 3.42±0.13 | 7 | 369.27±7.77 | 108.1±4.6 |
注:本次光释光测年所用矿物颗粒是粒径为 4-11 μm 的细颗粒石英。
6 总结
本文报道的产自河北蔚县南柏山晚更新世遗址的鹿角化石,是迄今发现的最完整的河套大角鹿鹿角标本,其主枝掌部最大长宽是670×526.8 mm,其眉枝的长宽是510×480 mm;角干最大长度和最大周长分别是270 mm和193 mm;角环最大周长是310 mm。河套大角鹿的鹿角掌部以其蒲扇状为特征,其边缘较为圆滑而不分叉,与肿骨鹿和扁角鹿及欧洲大角鹿的角区别明显。南柏山遗址的鹿角应当是从一成年河套大角鹿个体头上脱落的。该鹿角产自河流相沉积,含化石地层的光释光测年数据约为距今11万年前。
致谢
河北博物院郝建文研究员提供遗址线索;北京大学博士生马宁、河北师范大学硕士生徐英俊和本科生铁卫冬参加遗址发掘;铁卫冬协助光释光样品前处理;北京大学夏正楷教授、张家富教授指导岩性和沉积分析;贾真岩对化石进行现场加固和提取;河北省文物局、河北博物院驻村精准扶贫工作组、河北省文物考古研究院和蔚县文物局对野外工作给予大力支持;中国科学院古脊椎动物与古人类研究所标本中心李淳主任指导化石的室内处理;张伟工程师扫描标本并完成影像处理;丁今朝技师精心修理和加固标本;许勇先生绘制复原图;北京自然博物馆信息中心安萌老师在借阅图书资料时提供帮助;北京大学江左其杲博士和中国科学院古脊椎动物与古人类研究所白炜鹏博士分享有关文献;内蒙古博物院李虹女士提供对比化石信息;中国科学院南京地理与湖泊研究所侯彦冬博士在ArcGIS软件使用过程中提供帮助;与南京师范大学陈曦博士进行过有益探讨和交流;董为研究员和王世骐研究员审阅文稿并提出宝贵修改建议。作者对上述机构和个人表示衷心感谢!
谨以此文祝贺《人类学学报》创刊40周年!
参考文献
Morphology, taxonomy, and phylogeny of megacerines (Megacerini, Cervidae, Artiodactyla)
[J].DOI:10.1134/S0031030113080017 URL [本文引用: 16]
The phylogenetic position of the ‘giant deer’ Megaloceros giganteus
[J].DOI:10.1038/nature04134 [本文引用: 1]
The deer of the Weybourn Crag and forest bed of Norfolk
[J].
Phylogeny of the giant deer with palmate brow tines Megaloceros from west and Sinomegaceros from east Eurasia
[J].DOI:10.1016/j.quaint.2007.08.017 URL [本文引用: 8]
The evolution of the giant deer, Megaloceros giganteus (Blumenbach)
[J].DOI:10.1111/zoj.1994.112.issue-1-2 URL [本文引用: 3]
The first complete skeleton of Megaloceros verticornis (Dawkins, 1868) Cervidae, Mammalia, from Bilshausen (Lower Saxony, Germany): description and phylogenetic implications
[J].
Les Cervidés
In: Lavocat R. (Ed.). Faunes et flores préhistoriques de l’Europe occidental[M]. Éditions N. Boubée & C., Paris,
New fossils of Bison palaeosinensis (Artiodactyla, Mammalia) from the steppe mammoth site of Early Pleistocene in Nihewan Basin, China
[J].DOI:10.1016/j.quaint.2016.07.033 URL [本文引用: 1]
Skeleton of the giant deer Megaloceros giganteus giganteus (Blumenbach, 1803) (Mammalia, Artiodactyla) from the Irtysh Region near Pavlodar
[J].DOI:10.1134/S0031030114050104 URL [本文引用: 1]
Taxonomy, systematics and evolution of giant deer Megaloceros giganteus (Blumenbach, 1799) (Cervidae, Mammalia) from the Pleistocene of Eurasia
DOI:10.3390/quat4040036
URL
[本文引用: 2]

The article presents a preliminary morphological description of the holotype of Megaloceros giganteus (Blumenbach, 1799) that serves for the description of the species. The article proposes a taxonomical and morphological revision of the nominotypical subspecies M. giganteus giganteus and morphological comparison with other subspecies of M. giganteus. The cluster analysis of diagnostic craniodental and antler characters revealed the systematic position and phylogenetic relationships of M. giganteus with other cervid groups. The genus Praedama is regarded as a closely related phylogenetic branch that linked to the direct cursorial forerunner of Megaloceros that evolved in the middle latitudes of Western Siberia and northern Kazakhstan. The genus Dama has a distant relationship with Megaloceros and represents an earlier phylogenetic branch that evolved in the Ponto-Mediterranean area. The article discusses the secondary adaptations of M. giganteus forms to forest and woodland habitats in Europe and general paleobiogeographic features of the Megaloceros lineage.
Quaternary cave and fissure deposits and their fossils in Akiyosi District, Yamaguchi Prefecture
[J].
Giant deer: origin, evolution, role in biosphere
[J].DOI:10.1134/S0031030112070027 URL [本文引用: 1]
On the evolution of pachyostosis in jaw bones of Choukoutien giant deer Megaceros pachyosteus (Young)
[J].
New fossils of Eucladoceros boulei (Artiodactyla, Mammalia) from Early Pleistocene Nihewan Beds, China
[J].DOI:10.1016/j.palwor.2019.05.003 URL [本文引用: 1]
Mammalian fossils associated with the hominid skull cap of Lantian, Shensi
[J].
Antler morphology of the Yabe’s giant deer revision based on their ontogenetic variation
[J].
A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records
[J].
Bone and antler industry of the Choukoutien Sinanthropus site
[M].
Taphonomy at a distance: Zhoukoudian, “the cave home of Beijing Man?”
[J].DOI:10.1086/203303 URL [本文引用: 1]
Zhoukoudian: a closer look
[J].DOI:10.1086/203469 URL [本文引用: 1]
The Upper Cave fauna of Choukoutien
[M].
Les armes de chasse en matières osseuses dans la seconde moitié du Paléolithique récent. Douze millénaires d’histoire technique chez les chasseurs-collecteurs d’Europe occidentale (23000-11000 cal BP)
[J].
Giant deer Megaloceros giganteus Blumenbach, 1799 (Cervidae, Mammalia) from Palaeolithic of eastern Europe
[J].
A 51, 000-year-old engraved bone reveals Neanderthals’ capacity for symbolic behaviour
[J].
Stable isotope analysis of mammalian enamel from the Early Pleistocene site of Madigou, Nihewan Basin: Implications for reconstructing hominin paleoenvironmental adaptations in North China
[J].The reconstruction of environmental and climatic changes in the Pleistocene is an essential contribution to our understanding of human evolutionary and behavioral adaptations. Well preserved fluvio-lacustrine sediments at Nihewan basin have yielded a rich record of Early Pleistocene Paleolithic sites and mammalian fossils which provide a unique opportunity for exploring hominin behavior and paleoecology in North China. Taxonomic studies of mammalian fossils have provided important clues to the general environmental setting and landscapes of Early Pleistocene humans in the fluvio-lacustrine basin of Nihewan, but little is known about their isotopic signatures. In this paper, mammal teeth species at the Madigou archaeological site (ca. 1.2 Ma) were selected for bulk and sequential enamel stable isotope (C, O) analysis. Results show a variety of ecological environments, including grassland and sparse forest landscapes, and distinct patterns across taxa. C3-C4 mixed vegetation predominated, but C4 vegetation was also relevant at times. Madigou early humans likely experienced cold/warm or dry/wet fluctuations in this northern China basin. We hypothesize that the environmental fluctuations and diversified landscapes may have driven flexibility in various aspects of early human technological behaviors, and allowed hominins to face the environmental challenges of northern latitudes after the initial expansion from Africa into East Asia at the onset of the Middle Pleistocene Climate Transition.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}