


1 引言
2 地质背景
岩利1号洞位于广西壮族自治区崇左市大新县榄圩乡康合村西南约0.5 km处的岩利山,地理坐标为22°33′57″N、107°19′31″E,海拔约204 m。洞口朝东,海拔约204 m、宽约10 m、高约13 m(图1: a),洞道延伸长度约140 m。自2023年年底至2024年年初,对该洞穴进行了发掘,从内往外布三个探方(图1: b),分别为T1、T2和T3,三个探方的沉积序列相似。该洞共出土巨猿牙齿13枚,其中T3探方出土10枚、T1探方1枚(右p3,YLD1-043),地表清理2枚(左C,YLD1-SG-1;左m1/2,YLD1-SG-2)。以T3为例(探方深度为130 cm),自上而下可分为8层(图1: c),描述如下:
图1

图1
广西崇左岩利1号洞洞口照片(a)、平面图(b)及沉积剖面图(c)
Fig.1
The photograph(a), plane view(b) and deposition profile(c) of the Yanli Cave 1 in Chongzuo, Guangxi
1.表层钙板:厚5~10 cm。
2.黄褐色松散砂质黏土:含零星哺乳动物化石,厚0~15 cm。
3.钙板:厚5~10 cm。
4.浅褐色粉砂质黏土:部分钙质胶结,发育水平纹层,富含哺乳动物化石,厚0~15 cm。出土1枚巨猿牙齿(左P3,YLD1-589)。
5.红褐色粉砂质黏土:钙质胶结,质地坚硬,富含哺乳动物化石,为巨猿化石的主要产出层位,厚10~15 cm。出土巨猿牙齿7枚(左P3, YLD1-447;左M1/2, YLD1-418;右i2, YLD1-477;左p4, YLD1-653;右p4, YLD1-612;左m1/2, YLD1-326;左m, YLD1-Y01)。
6.浅褐色粉砂质黏土:钙质胶结,含哺乳动物化石,厚20~30 cm。出土巨猿牙齿1枚(左M1/2,YLD1-715)。
7.深褐色砂质黏土:钙质胶结,含零星哺乳动物化石,厚20~30 cm。出土巨猿牙齿1枚(右C,YLD1-416)。
8.浅褐色砂质黏土: 钙质胶结,质地坚硬,未见化石,厚20~30 cm,未见底。
3 研究方法
牙齿形态描述主要参照王頠与张颖奇等所使用的巨猿牙齿解剖术语体系[2,32]。采用游标卡尺对牙齿尺寸进行测量,精度为0.1 mm。齿冠近远中径(mesiodistal length, lMD)与颊舌径(buccolingual breadth, bBL)的测量参照White和Suwa等[33,34]。对于门齿、第四前臼齿和臼齿,lMD为齿冠近-远中平分轴的长度;对于犬齿和下颌第三前臼齿,lMD为平行于近-远中轴测得的最大长度;对于上颌第三前臼齿,lMD为近-远中之间的最大长度(该方向不一定会平行近-远中轴)。确定lMD后,bBL即为垂直于lMD方向上的最大宽度。为了行文简洁,后文所述长与宽分别指lMD和bBL。我们对其中1颗牙齿(YLD1-653)的尺寸测量进行了观察者内和观察者间误差分析(intra- and inter-observer error analysis),误差计算方式为:(最大值-最小值)/最小值。结果显示,长和宽的观察者内误差与观察者间误差均小于5%。
4 化石描述
4.1 上颌牙齿
上颌犬齿(C):2枚,YLD1-416和YLD1-SG-1。YLD1-416,长16.0 mm、宽15.1 mm (图2: a),齿冠完整,齿根仅存残留,其表面可见啮齿类啃咬痕迹。该牙齿较为粗壮,齿冠相对较低,呈颊舌向压缩的圆锥形。近中缘钝圆,伴生纵向浅沟,远中缘较锐利。舌面前半稍隆起,后半相对低凹并发育明显的纵向指状突。舌面基部可见典型的V型齿带,颊面基部远中侧残留齿带痕迹。齿尖可见倾向远中侧的磨耗面,齿质暴露,剖面显示釉质层较厚。YLD1-SG-1,长16.3 mm、宽15.6 mm(图2: b),整体形态与YLD1-416相似,不同点在于:前者磨耗程度更大,齿质大面积暴露;未见颊面齿带残留。
图2
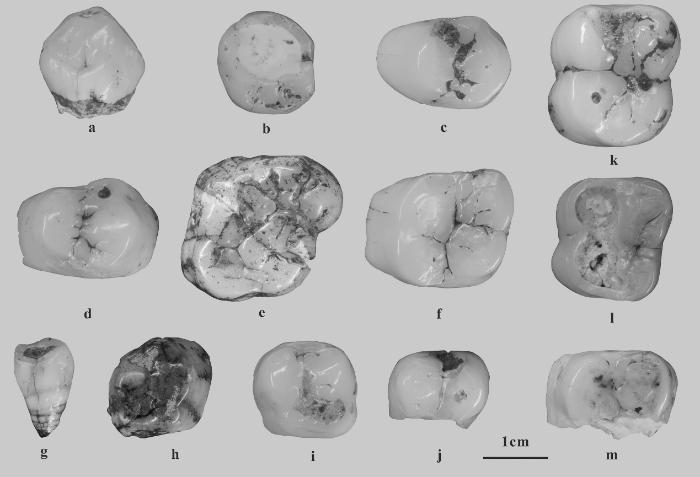
图2
岩利1号洞出土的巨猿化石
a. 右right C, YLD1-416;b. 左left C, YLD1-SG-1;c. 左left P3, YLD1-447;d. 左left P3, YLD1-589;e. 左left M1/2, YLD1-715;f. 左left M1/2, YLD1-418;g. 右right i2, YLD1-477;h. 右right p3, YLD1-043;i. 左left p4, YLD1-653;j. 右right p4, YLD1-612;k. 左left m1/2, YLD1-326;l. 左left m1/2, YLD1-SG-2;m. 左left m, YLD1-Y01
Fig.2
The Gigantopithecus fossils excavated from Yanli Cave 1
上颌第三前臼齿(P3):2枚,YLD1-447和YLD1-589。YLD1-447,长14.2 mm、宽20.1 mm,齿冠完整,齿根局部残留(图2: c)。标本呈粗壮高冠形态,宽大于长,周缘无齿带。咬合面呈三角形(颊侧部分最长,并向舌侧逐渐收窄)。中央纵沟沿不规则路径贯穿咬合面,形成不对称的颊舌侧分区(颊侧面积占比更大)。前尖与原尖通过未分叉的横脊相连,前尖在体积与高度上均显优势。近中凹和远中凹均表现为横向细缝,其中远中凹发育更好,且其两侧各附有一个结节。近中面颊侧具明显的瘤状突起,远中面舌侧可见断续纵沟。YLD1-589,长15.5 mm、宽21.7 mm(图2: d)。相比YLD1-447,YLD1-589的横脊在中央凹处被三条横向细沟分割。
上颌第一/二臼齿(M1/2):2枚,YLD1-715和YLD1-418。YLD1-715,长22.1 mm、宽25.6 mm,齿冠高耸,齿根仅存基部(图2: e)。咬合面呈菱形轮廓,宽大于长,颊舌向收腰(buccolingual waisting)不明显。沿着远中方向,牙齿稍微变窄。齿尖硕大圆钝,并挤满咬合面,导致齿凹的辨识度较低。四个齿尖发育完整,其中原尖、前尖和后尖大小相近,且均大于次尖。齿尖高度序列为:前尖>原尖≈后尖>次尖。各齿尖被深邃的细沟分隔,前尖较原尖明显向前突出,而次尖比后尖明显向后突出。齿带缺失。近中面和远中面上均有椭圆形齿间接触面。YLD1-418,长16.8 mm、宽21.8 mm(图2: f)。相比YLD1-715,YLD1-418的整体尺寸更小,各个齿尖也更小,咬合面近似矩形;前尖和后尖近似等高,高于原尖和次尖;原尖与次尖的突出程度减小。
4.2 下颌牙齿
下颌侧门齿(i2):1枚,YLD1-477,长8.9 mm、宽11.1 mm,齿冠完整,齿根仅存基部(图2: g)。齿冠低,呈钉状。切缘与近中面构成将近90度的近中角,与远中面则形成大于90度的远中角。切缘磨耗面呈椭圆形,暴露出透镜状齿质,其中近中部齿质暴露宽度显著大于远中部。切缘唇侧可见崩裂疤痕(enamel chipping)。舌侧面结构特征中,远中残留齿带,近中齿带缺失,因而缺少典型的“V”形齿带结构;釉质表面整体较为光滑,未见明显褶皱和指状突。
下颌第三前臼齿(p3):1枚,YLD1-043,长16.4 mm、宽14.4 mm(图2: h)。齿冠完整,齿根仅存基部。长大于宽,但与现生大猿同齿位相比,其近远中径相对较短而牙齿整体显得更为紧凑。齿冠相对较高,近中颊侧面发育高耸陡峭的研磨面。咬合面轮廓呈扇形,近中颊侧见瘤状凸起。咬合面具有两个发育完整且呈横向排列的主齿尖。其中下原尖较下后尖更为硕大,但因磨耗致齿质呈点状暴露,进而高度更低。两齿尖紧密相邻,被一条清晰的中央纵沟分隔。近中凹呈坑状,远中凹呈横向深沟状,二者均发育良好并被中央纵沟连接。近中边缘脊和远中边缘脊发育良好,其中远中边缘脊更为圆钝。远中边缘脊与下原尖后脊交汇处可见两个小型附属结节。
下颌第四前臼齿(p4):2枚,分别为YLD1-653与YLD1-612。YLD1-653,长14.1 mm、宽15.3 mm,齿冠高耸完整,齿根保留较多但仍不完整(图2: i)。齿冠较高,该齿呈现典型臼齿化特征,具有两个主齿尖和一对远中结节。咬合面轮廓近似方形,宽略大于长。下原尖在体积上略占优势,但因磨耗致使其高度略低于下后尖。两个主齿尖间距宽阔,由一条发育良好的横脊相连,该横脊被中央纵沟贯穿分割。近中凹表现为横向浅沟状,近中边缘脊纤细且表面起伏不平。近中面和远中面均可见椭圆形齿间接触面。
YLD1-612标本宽15.5 mm,两个主齿尖保存较为完整,但齿冠远中部分缺失,且齿颈线存在破损(图2: j)。从断裂面可观察到很厚的釉质层。该牙齿为高冠形态。由于磨耗作用,下原尖的高度低于下后尖,并出现点状齿质暴露。近中凹因磨耗改造而呈现为裂隙状。此外,在近中面上观察到龋窝。
下颌第一/二臼齿(m1/2):2枚,YLD1-326和YLD1-SG-2。YLD1-326,长21.6 mm、宽19.0 mm,齿冠较为完整(基部稍有缺损),齿根近乎全部缺失(图2: k)。该齿为高冠型,尺寸硕大,长大于宽,宽度向远中方向略收缩。颊、舌侧面中部均发育显著纵向沟,颊舌向收腰特征明显。颊侧残留齿带痕迹。咬合面呈矩形,五个齿尖发育良好,其大小关系为:下后尖>下原尖≈下次尖>下内尖≈下次小尖。磨耗导致齿尖高度分异:下后尖最高,下原尖次之,下次尖、下内尖与下次小尖最低。连接下原尖与下后尖的横脊粗壮圆钝,被中央纵沟隔断。下内尖与下后尖之间可见一个小型结节。近中凹表现为横向裂隙状,近中边缘脊纤细且附一小结节。近、远中面均具椭圆形齿间接触面。YLD1-SG-2,长19.1 mm、宽17.2 mm(图2: l)。相比YLD1-326,YLD1-SG-2的整体尺寸更小、颊侧磨耗严重,所以无法观察下原尖、下次尖和下次小尖的形态;颊侧未见齿带残留;下后尖和下内尖之间的小结节尺寸更大;近中面上有两个齿间接触面。
下颌臼齿(m)还有一枚左侧齿,YLD1-Y01,宽20.4 mm(图2: m)。该牙齿为高冠型,仅保存近中部分。下原尖和下后尖尺寸相近,但下原尖因磨耗高度较低,两尖之间可见明显的纵沟。在牙齿断裂面处可观察到很厚的釉质层。标本远中部位缺失,难以判定具体齿位。
5 讨论
5.1 岩利1号洞哺乳动物群的年代
岩利1号洞出土的哺乳动物化石组合共鉴定出6目17科27属30种。灵长目包括步氏巨猿(G. blacki)、魏氏猩猩(Pongo weidenreichi)、长臂猿属未定种(Hylobates sp.)、猕猴属未定种(Macaca sp.)、金丝猴属未定种(Rhinopithecus sp.)及乌叶猴属未定种(Trachypithecus sp.)。啮齿目包括江山豪猪(Hystrix kiangsenensis)。食肉目包括古爪哇豺(Cuon javanicus antiquus)、虎(Panthera tigris)、云豹(Neofelis nebulosa)、金猫(Catopuma temminckii)、大灵猫属未定种(Viverra sp.)、亚洲黑熊(Ursus thibetamus)、武陵山大熊猫(Ailuropoda wulingshanensis,图3: a-b)、巴氏大熊猫(A. melanoleuca baconi,图3: c)、南方猪獾(Arctonyx collaris)与亚洲狗獾(Meles leucurus)。奇蹄目包括爪哇犀(Rhinoceros sondaicus,图3: d)与中国貘(Tapirus sinensis,图3: e)。偶蹄目则有裴氏猪(Sus peii)、小猪(S. xiaozhu)、水鹿(Cervus unicolour)、毛冠鹿(Elaphodus cephalophus)、赤麂(Muntiacus muntjak)、小麂(M. reevesi)、苏门鬣羚(Capricornis sumatraensis)、广西巨羊(Megalovis guangxiensis,图3: f)与大额牛(Bos gaurus)。长鼻目含东方剑齿象疑似种(Stegodon cf. S. orientalis,图3: g)与亚洲象(Elephas maximus,图3: h)。
图3
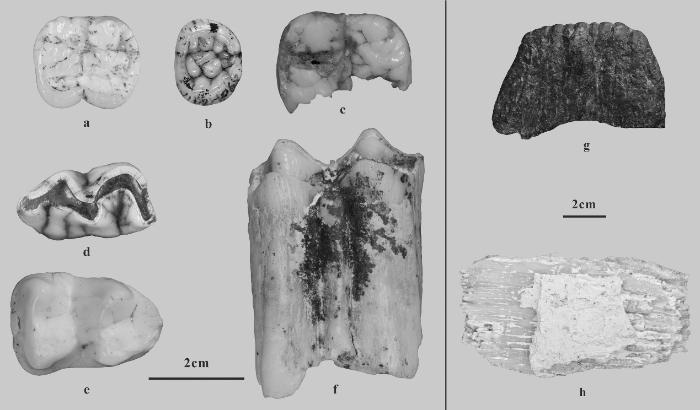
图3
岩利1号洞出土的部分哺乳动物化石
a.武陵山大熊猫/A. wulingshanensis;b.武陵山大熊猫/A. wulingshanensis;c.巴氏大熊猫/A. melanoleuca baconi;d.爪哇犀/R. sondaicus;e.中国貘/T. sinensis;f.广西巨羊/M. guangxiensis;g.东方剑齿象疑似种/Stegodon cf. S. orientalis;h.亚洲象/E. maximus。除了亚洲象发现于地表扰乱层之外,图中其他属种均可见于含巨猿化石的地层Apart from the E. maximus, which was found in a surface - disturbed layer, all other taxa shown in the figure can be found in the strata containing G. blacki
Fig.3
Some mammalian fossils excavated from Yanli Cave 1
岩利1号洞的地质年代为早更新世早期的可能性较低,因为动物群中缺失该时期典型物种,如大熊猫小种(A. microta)与中华黄昏爪兽(Hesperotherium sinense)。该洞穴与缺缺洞的动物群组成呈现较高相似性,二者都有步氏巨猿、广西巨羊、中国貘及巴氏大熊猫等物种(图3)。同时,两处洞穴均位于崇左地区,海拔相近(约200 m),表明其地质年代可能接近。然而,两地动物群的差异也值得关注:缺缺洞出土早更新世典型物种中华乳齿象(Sinomastodon sp.),而岩利1号洞未发现该属种,但可能存在早更新世晚期才首现的东方剑齿象(图3: g)。这一对比指示岩利1号洞的年代可能晚于缺缺洞。缺缺洞已获得的直接测年结果为1151±332 ka至704±150 ka,推定年代约为1.0 Ma[9,21],故岩利1号洞年代可能晚于1.0 Ma。再者,岩利1号洞同时保存有早更新世晚期典型物种武陵山大熊猫与中更新世典型物种巴氏大熊猫(图3: a-c)。总的来说,岩利1号洞动物群兼具早更新世晚期与中更新世典型元素,且年代可能晚于缺缺洞,呈现出早-中更新世过渡期特性。不过,要确定最终的精确年代,还需要依靠其他直接测年手段,例如古地磁和铀系测年。
5.2 与其他巨猿遗址的牙齿尺寸对比
图4
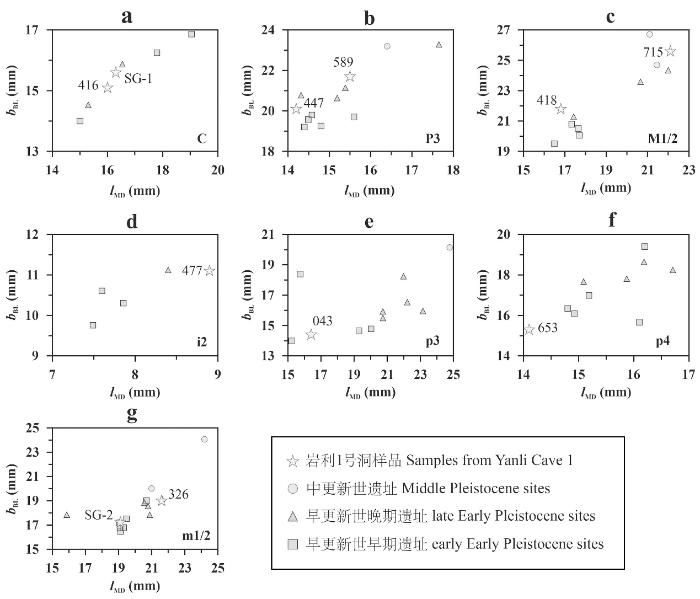
图4
岩利1号洞和其他巨猿遗址牙齿尺寸的对比
a.上颌犬齿Upper canine;b.上颌第三前臼齿Upper third premolar;c.上颌第一/二臼齿Upper first/second molar;d.下颌侧门齿Lower lateral incisor;e.下颌第三前臼齿Lower third premolar;f.下颌第四前臼齿Lower fourth premolar;g.下颌第一/二臼齿Lower first/second molar。图中其他遗址的巨猿牙齿尺寸均为平均值,数据来源于文献[11,13,16,19,29,30,35] / Data points for other sites represent mean Gigantopithecus tooth dimensions, derived from references [11,13,16,19,29,30,35]
Fig.4
Dental size comparison between Yanli Cave 1 and other Gigantopithecus sites
总的来看,岩利1号洞巨猿的前齿尺寸中等偏大,而在颊齿中,既有一部分标本呈现出早更新世早期的较小尺寸特征(447、418、043、653、SG-2),但也有标本接近早更新世晚期尺寸(589和326),甚至接近中更新世尺寸(715)。这一现象暗示在早-中更新世过渡阶段,岩利1号洞种群可能出现了颊齿尺寸的适应性增大趋势。然而,此初步认识仍存在若干局限。首先,作为猩猩亚科成员,巨猿可能与其现生近亲猩猩相似,在前臼齿尺寸上存在两性异形现象(sexual dimorphism)[36,37]。若该推测成立,则前臼齿尺寸可能形成双峰分布模式,通常表现为雌性小于雄性,从而在一定程度上干扰了对前臼齿整体演化趋势的认识。其次,受当前样本数量所限,现有结论尚未经过统计学检验,其可靠性仍需更多材料支撑。
6 结论
广西崇左岩利1号洞出土的巨猿化石,在形态特征与尺寸上和已知巨猿相似。伴生动物群指示其年代可能处于早-中更新世过渡期,然精确年代需借助直接定年技术予以确认。该遗址巨猿牙齿既有偏小尺寸(接近早更新世早期巨猿),也有偏大尺寸(接近早更新世晚期-中更新世巨猿),在一定程度上显示出早-中更新世过渡期的特性。然此初步认识存在一定局限,尤以样本数量不足为甚。未来若能综合对比岩利1号洞、缺缺洞与展望洞这三处年代接近早-中更新世过渡期的巨猿遗址材料,将有望揭示气候转型背景下巨猿牙齿性状的演化模式与适应策略。
致谢
感谢广西自然博物馆、广西民族博物馆、南宁师范大学、崇左市大新县康合村干部及村民在洞穴发掘过程中给予的支持和帮助。感谢中国科学院古脊椎动物与古人类研究所张颖奇、邢松和张立召三位老师在巨猿牙齿观察对比上的帮助。
参考文献
Eine fossile Säugetierfauna mit Simiaaus Südchina
[J].
Gigantopithecus blacki: A giant ape from the Pleistocene of Asia revisited
[J].
Giant early man from Java and South China
[J].
Gigantopithecus
[J].
巨猿牙齿釉质厚度及对食性适应与系统演化的意义
[J].
Enamel proteome shows that Gigantopithecus was an early diverging pongine
[J].
The demise of the giant ape Gigantopithecus blacki
[J].
New discoveries of Gigantopithecus blacki teeth from Chuifeng Cave in the Bubing Basin, Guangxi, South China
[J].
广西崇左三合大洞新发现的巨猿动物群及其性质
[J].
A fourth mandible and associated dental remains of Gigantopithecus blacki from the Early Pleistocene Yanliang Cave, Fusui, Guangxi, South China
[J].
Giant primate of Java: A new Gigantopithecus specimen from Semedo
[J].
广西布兵盆地么会洞发现的巨猿牙齿化石
[J].
Chronological sequence of the early Pleistocene Gigantopithecus faunas from cave sites in the Chongzuo, Zuojiang River area, South China
[J].
New 400-320 ka Gigantopithecus blacki remains from Hejiang Cave, Chongzuo City, Guangxi, South China
[J].
The middle Pleistocene transition: Characteristics, mechanisms, and implications for long-term changes in atmospheric pCO2
[J].
A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records
[J].
中更新世气候转型:特征、机制和展望
[J].
Northern hemisphere ice sheet expansion intensified Asian aridification and the winter monsoon across the mid-Pleistocene transition
[J].
The Mid-Pleistocene Climate Transition
[J].
Variability and evolutionary trends in tooth size of Gigantopithecus blacki
[J].
Evolutionary trend in dental size in Gigantopithecus blacki revisited
[J].
Possible change in dental morphology in Gigantopithecus blacki just prior to its extinction: Evidence from the upper premolar enamel-dentine junction
[J].
New fossil hominids from Laetolil, Tanzania
[J].
Paleobiological implications of the Ardipithecus ramidus dentition
[J].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}